Страница: 5/9
Измерение пройденного пути
Наиболее естественный показатель движения клетки в эксперименте, считается пройденный путь. На рис. 12 можно наблюдать нарастание пройденного пути в зависимости от времени.
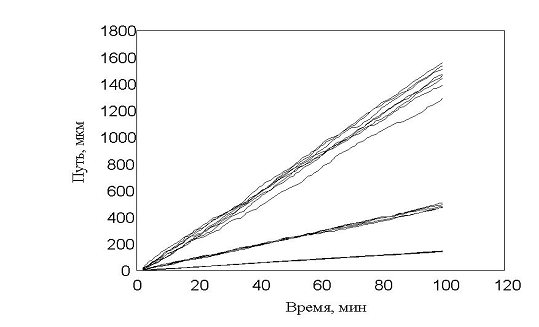
рис. 12 Зависимость пройденного пути от времени. Три группы клеток (max_step: 3, 10, 30 mcm - соотношение 15:30:55)
Показано движение 15 клеток из 30, которые генерировала модель. Наиболее удивительным в эксперименте был факт постоянной скорости движения клеток в течении всего эксперимента, не смотря на случайный характер их движения. Модель хорошо воспроизводит этот факт, при этом случайные колебания скорости практически незаметны. Каждая отдельная кривая представленная на рис. 12 хорошо соответствует экспериментальным данным рис. 13. Однако из сравнения рис. 12 и рис. 13 видно, что в эксперименте существуют все варианты скоростей движения, а в модельном эксперименте, четко выделяются три группы клеток.
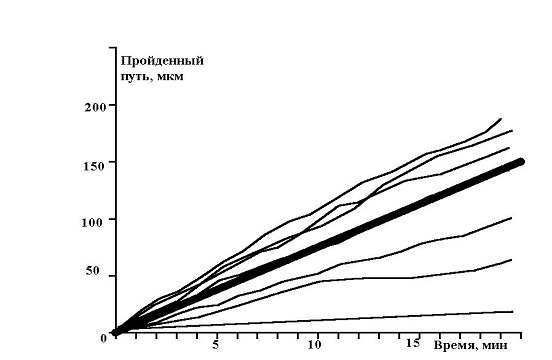
рис. 13 Зависимость пройденного пути от времени (эксперимент)
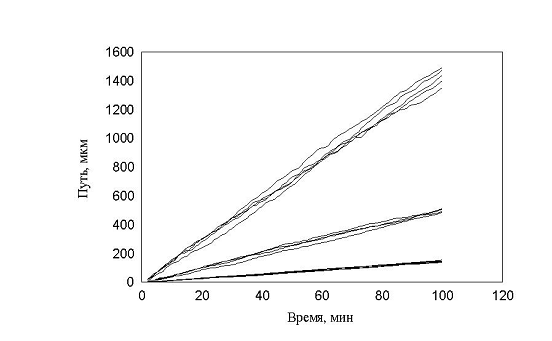
рис. 14 Зависимость пройденного пути от времени. Три группы клеток (max_step: 3, 10, 30 mcm соотношение 1:1:1)
Таким образом, условное деление клеток на группы, введенное экспериментатором не может быть основой для моделирования. При изменении процентного состава клеток в популяции рис. 14 видно, что характер распределения клеток по скоростям не изменяется. В обоих случаях четко выделяются три группы клеток, которые и были заложены в модель.
При моделировании популяции был проведен еще один эксперимент рис. 15, все параметры движения были такими, как и в предыдущем эксперименте.
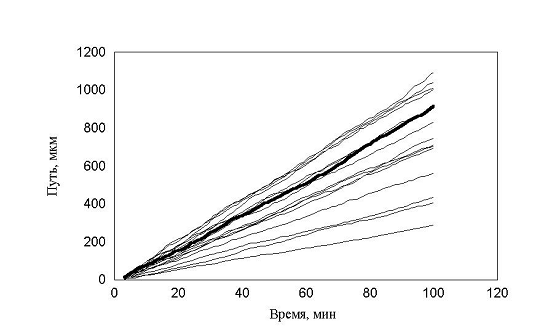
рис. 15 Зависимость пройденного пути от времени. Три группы клеток (max_step: 3, 10, 30 mcm, соотношение 1:1:1 время измерения каждый десятый шаг)
Однако «теоретический наблюдатель» имел возможность регистрировать положение клеток только на каждом 10-м шаге. В этом случае существование трех групп клеток практически не видно. Это связано с тем, что траектория движения клеток запутанная. Часто встречаются случае, когда клетки за 10 шагов успевают повернуть на зад. Таким образом, на каких-то участках траектории «возникает иллюзия» малых шагов у быстрых клеток и возникает картина, похожая на экспериментальную. Интересно отметить, что максимальная скорость измерения упала в 3 раза (приблизительно Ö10) по сравнению с заданной в программе. Последний расчет указывает на то, что выбор интервала времени, для последовательного положения клеток, очень существенен.
В экспериментах положение клетки оценивалось как положение её центра масс. Ясно, что координаты меняются от изменения формы клеток. Из этого следует, что чем меньше интервал времени, между измерениями, тем точнее измерение скорости. Однако смещение центра масс на расстояние меньше размера клеток нельзя считать устойчивым. В результате, для измерения скорости был выбран интервал 1 минута, который примерно соответствует смещению средних клеток на расстояние порядка их размера.
Последний рисунок не смотря с его схожестью с экспериментом не может считаться истинным. Таким образом приходим к выводу, что деление сделанное экспериментатором на быстрые, средние и медленные клетки очень условно и его можно принимать только как некую мнемонику.
Таким образом, было введено принципиальное изменение в модели рис. 16. В популяции присутствовали клетки с непрерывно распределенными скоростями, а не три ярко выраженные группы. Распределение минутных сдвигов для каждой клетки было принципиально таким же, как и в предыдущей модели. Сдвиги распределены от 0 до rmax, а величины rmax в популяции были распределены в соответствии с таким же распределением. Генерировалась случайная популяция клеток, после чего для каждой клетки генерировались сдвиги, и рассчитывалось движение клеток в популяции. Результат расчета, показанный на рис. 16, полностью совпадает с экспериментальным. Мы моделировали не только перемещение, но и процесс измерения перемещений. Процедура измерений перемещений просматривала все движущиеся клетки, не зная к какой группе относится каждая из них.
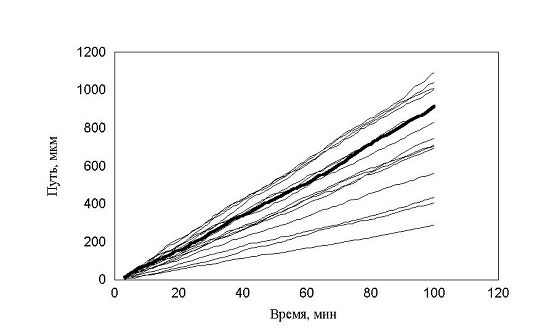
рис. 16 Зависимость пройденного пути от времени. Клетки распределены непрерывно (max_step: 30 mcm, соотношение случайное, время измерения каждый шаг)
В результате она смешивала все клетки и строила гистограмму
рис. 17 – 19. Все полученные гистограммы не имели характерных особенностей и
были, в принципе, похожи друг на друга. В результате приходим к важному выводу
– измерение гистограмм, в эксперименте, не дает существенной информации о характере
движения. Такой же вывод будет сделан и в параллельной работе при более детальном
анализе каждой отдельной клетки.
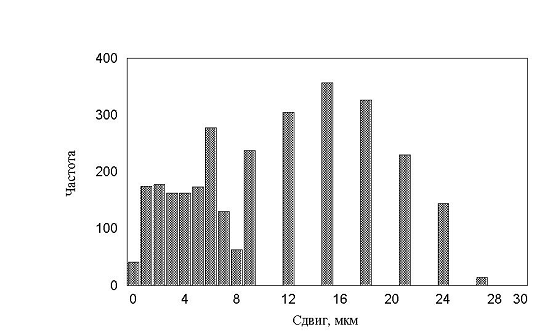
рис. 17 Гистограмма минутных сдвигов. Три группы клеток (max_step 3, 10, 30 соотношение клеток 15:30:55)
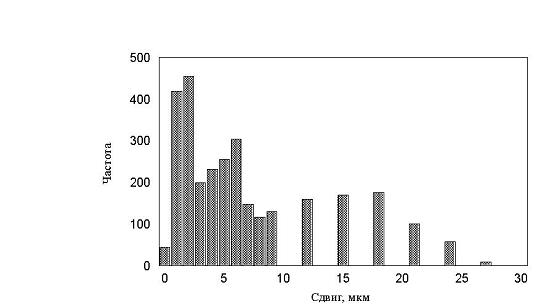
рис. 18 Гистограмма минутных сдвигов. Три группы клеток (max_step 3, 10, 30 соотношение клеток 1:1:1)
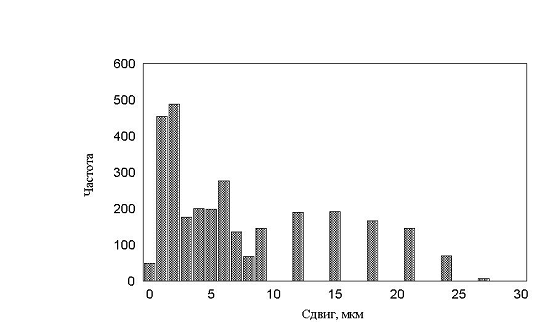
рис. 19 Гистограмма минутных сдвигов. Три группы клеток (max_step 3, 10, 30 соотношение клеток 1:1:1, время измерения каждый десятый шаг)
Исследование зависимости среднеквадратичного отклонения от времени наблюдения
Как известно из теории Броуновского движения, важным показателем случайного блуждания является среднеквадратичное отклонение. Мы использовали этот показатель для движения нейтрофилов. Прежде всего, мы рассматривали частицу, у которой углы поворотов распределены равномерно, т.е. клетка с равной вероятностью может повернуть в любую сторону. Результат показан на рис. 20.
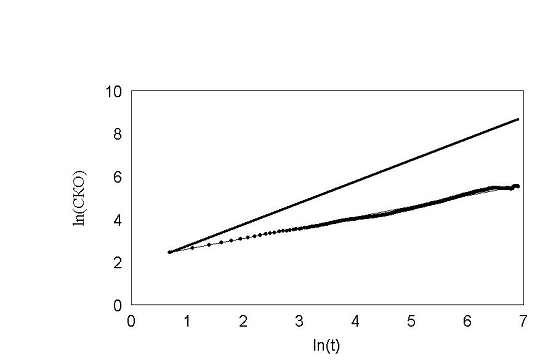
рис. 20 Расномерное распределение углов.
На этом рисунке по оси ординат отложен ln времени. По оси абсцисс отложен ln среднеквадратичного отклонения клетки. Как показано Энштейном и Смолуховским для броуновской частицы, эта зависимость отображается прямой с наклоном 0,5. Как видно, наша кривая хорошо ложится на прямую с наклоном 0,5. Для сравнения, на этом же графике проведена прямая с наклоном 1. Если бы клетка не поворачивалась вообще, то её среднеквадратичное отклонение легло бы на прямую с наклоном 1. Результат показан на рис. 21.
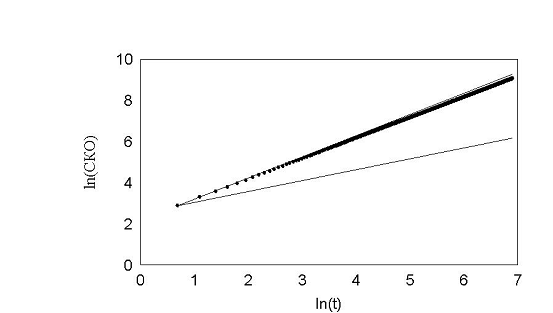
рис. 21 Почти нулевой угол поворотов.
Здесь на всякий случай оставили небольшую вероятность (2%) поворота на 180. В результате среднеквадратичное отклонение этих клеток слегка отклоняется от прямой.
При задании в модели экспериментально измеренных распределений углов поворотов рис. 22 получили кривые поразительно похожие на эксперимент. В начале, среднеквадратичное отклонение идет вдоль прямой с наклоном 1, затем отклоняется и асимптотически стремиться к наклону 0,5 рис. 23 – 24. Такой характер зависимости в литературе принято трактовать как клеточную память. В нашей модели процесс был принципиально марковским. В каждый момент времени распределение углов и сдвигов не зависело от предыстории. И тем не менее картина воспроизводилась с точностью до времен. Конечно если трактовать память в бытовом смысле, то элементы памяти в модели есть. Все-таки нулевой угол поворота по отношению к предыдущему шагу является предпочтительным.
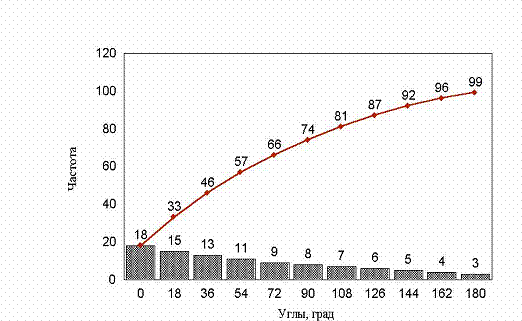
рис. 22 Гистограммы углов заданных поворотов.
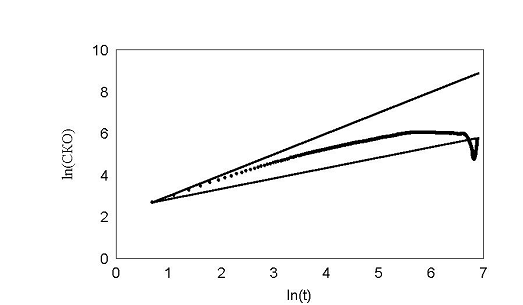
рис. 23 Обычное распределение углов fast.
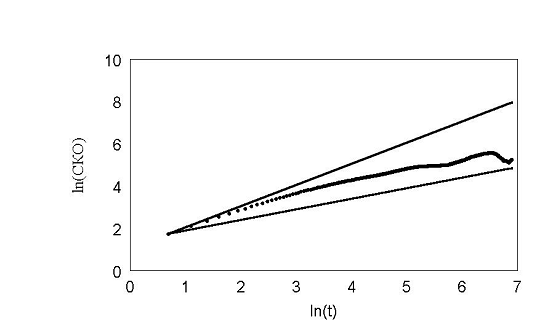
рис. 24 Обычное распределение углов slow.
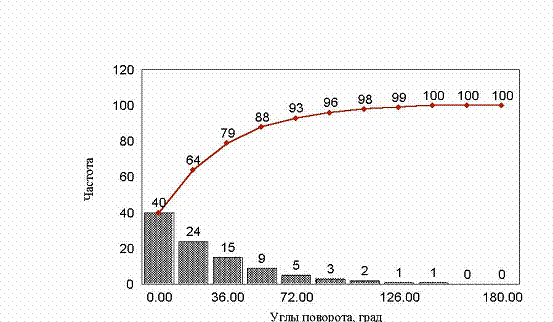
рис. 25 Гистограмма углов заданных поворотов (хемотаксис)
Однако памяти, в математическом смысле, в модели нет.
Если мы сделаем нулевой угол еще более предпочтительным рис. 25, то естественно получим кривую среднеквадратичного отклонения, очень близкую к кривой, получаемой при хемотаксисе.
Реферат опубликован: 23/05/2005 (33137 прочтено)