Страница: 3/7
Белки, образующие цитоскелет эритроцитов, неуникальны. Они или им подобные белки есть во многих других клетках. Преимущественная их локализация – вблизи мембран. Вероятно также, что рассмотренные белки могут быть полифункциональны и использоваться в структурных комплексах разного функционального назначения. [5]
Полосу 9 на электрофореграмме составляет белок глобин, который является частью мембрансвязанного гемоглобина. Как уже говорилось выше, цитоплазматический домен белка полосы 3, включающий около 380 аминокислотных остатков, локализован в цитоплазме и играет важную роль в поддержании формы эритроцита и его метаболизме. N-концевая последовательность цитоплазматического домена богата аминокислотными остатками кислого характера. На этом участке находятся центры связывания гидролитических ферментов – глицеральдегид-3-фосфатдегидрогеназы, альдолазы, каталазы, гемоглобина и гемихрома. Таким образом, цитоплазматический домен анионтранспортного белка и является местом локализации мембрансвязанного гемоглобина, о структуре, свойствах и некоторых аспектах функционирования которого будет сказано ниже.
Гемоглобин Гемоглобин – белок эритроцитов, красных кровяных клеток, переносящий молекулярный кислород от легких к тканям в организмах позвоночных животных. Гемоглобин можно считать своего рода модельным белком, структура, свойства и функции которого наиболее полно изучены по сравнению с другими белками на протяжении последних 50 лет. Американский физик Хопфилд назвал его атомом водорода современной биохимии, имея в виду, что изучение гемоглобина сыграло в биохимии ту же роль, что и изучение атома водорода в физике. Гемоглобин называют также почетным ферментом, поскольку исследования его структуры в статике и динамике позволили значительно продвинуться в понимании механизмов функционирования ферментов. Структура этого глобулярного белка известна в деталях главным образом благодаря работам английского биофизика Макса Перутца, который получил первые рентгеноструктурные данные еще в конце 40-х годов нашего века. За эти исследования он был удостоен Нобелевской премии.
Структура гемоглобина Молекула гемоглобина состоит из четырех субъединиц: двух a и двух b - и соответственно содержит четыре полипептидные цепочки двух сортов. Каждая a-цепочка содержит 141, а b-цепочка – 146 аминокислотных остатков. Таким образом, вся молекула гемоглобина включает 574 аминокислоты. Хотя аминокислотные последовательности a- и b-цепочек различны, они имеют практически одинаковые третичные пространственные структуры. Собственно говоря, приведенные выше детали структуры относятся не к гемоглобину, а к его белковой компоненте – глобину. Каждая субъединица гемоглобина содержит одну небелковую (так назваемую простетическую) группу – гем. Гем представляет собой комплекс Fe(II) с протопорфирином. Структура гема представлена на рис.1, а. [2]
Атом железа может образовать шесть координационных связей. Четыре связи направлены к атомам азота пиррольных колец, оставшиеся две связи –
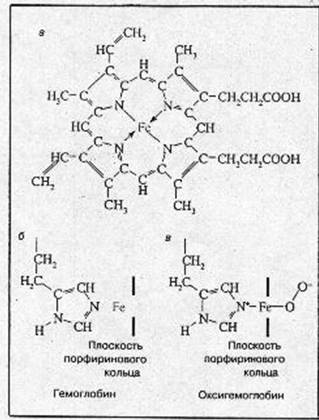
Рис.1. Структура гема (а), структура активного центра дезоксигемоглобина (б), структура активного центра оксигемоглобина (в).
перпендикулярно к плоскости порфиринового кольца по обе его стороны. Гемы расположены вблизи поверхности белковой глобулы в специальных карманах, образованных складками полипептидных цепочек глобина. Гемоглобин при нормальном функционировании может находиться в одной из трех форм: феррогемоглобин (обычно называемый дезоксигемоглобином или просто гемоглобином), оксигемоглобин и ферригемоглобин (называемый также метгемоглобином). В феррогемоглобине железо находится в закисной форме Fe(II), одна из двух связей, перпендикулярных к плоскости порфиринового кольца, направлена к атому азота гистидинового остатка, а вторая связь свободна (рис.1, б). Кроме этого гистидинового остатка, называемого проксимальным (соседним), по другую сторону порфиринового кольца и на большем расстоянии от него находится другой гистидиновый остаток — дистальный гистидин, не связанный непосредственно с атомом железа. Взаимодействие молекулярного кислорода со свободным гемом приводит к необратимому окислению атома железа гема [Fe(II) Þ Fе(III); гем Þ гемин]. В дезоксигемоглобине глобин предохраняет железо гема от окисления.
Реакция оксигенации Обратимое присоединение кислорода (оксигенация), позволяющее гемоглобину выполнять свою основную функцию переносчика, обеспечивается возможностью образовать прочные пятую и шестую координационные связи и перенести электрон на кислород не от железа (то есть окислить Fe2+), а от имидазольного кольца проксимального гистидина. Это схематически изображено на рис.1, б. Вместо молекулярного кислорода железо гема может присоединить окись углерода СО (угарный газ). Даже небольшие концентрации СО приводят к нарушению кислородпереносящей функции гемоглобина и отравлению угарным газом.
Выше было сказано, что одна молекула гемоглобина содержит четыре субъединицы и, следовательно четыре гема, каждый из которых может обратимо присоединить одну молекулу кислорода. Поэтому реакцию оксигенации можно разделить на четыре стадии:
Hb+O2 Û HbO2 (1a)
НbO2+O2 Û Hb(O2)2 (1б)
Hb(O2)2+O2 Û Hb(O2)3 (1в)
Hb(O2)3+O2 Û Hb(O2)4 (1г)
Прежде чем рассмотреть эту главную функциональную реакцию гемоглобина более детально, необходимо сказать несколько слов о мышечном гемоглобине — миоглобине. Этот красящий белок поперечнополосатых мышц представляет собой комплекс гема с "четвертушкой" глобина. Он содержит одну молекулу гема и одну полипептидную цепочку, состав и структура которой подобны составу и структуре b-субъединицы гемоглобина. Как и для гемоглобина, важнейшей функцией миоглобина является обратимое присоединение молекулярного кислорода. Эту функцию характеризует так называемая кривая оксигенации, связывающая степень насыщения гемоглобина кислородом (в процентах) с парциальным давлением последнего, рО2 (мм Hg). Типичные кривые оксигенации гемоглобина и миоглобина (при условии достижения химического равновесия) приведены на рис.2, а, б. Для миоглобина кривая является гиперболой, как и должно быть в случае одностадийной химической реакции при условии достижения химического равновесия:
Mb+O2ÛMbO2 (2)
где Mb - миоглобин.
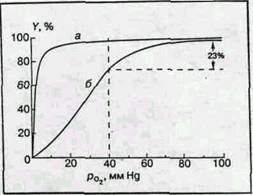
Рис. 2 Кривые оксигенации
миоглобина (а) и гемоглобина (б)
Совершенно другая картина возникает в случае гемоглобина. Кривая диссоциации имеет S-образную форму. Без кислорода молекулы гемоглобина обладают низким сродством к кислороду и равновесие реакции (1а) сдвинуто влево. Затем кривая становится круче и при высоких значениях рО2 практически сливается с кривой диссоциации миоглобина. [2]
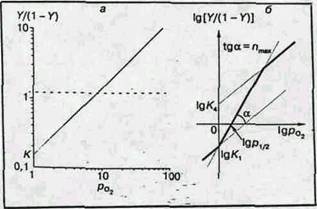
Рис. 3. Логарифмические анаморфозы кривых оксигенации миоглобина (a) и гемоглобина (б)
М.Перутц пишет, что распределение молекул кислорода по молекулам гемоглобина следует библейской притче: "Каждому, у кого есть, дай еще, и у него будет избыток; у того же, у кого нет, забери то немногое, что у него осталось". Это заставляет предположить, что между гемами одной молекулы гемоглобина существует некоторая связь, благодаря которой присоединение кислорода к одному гему влияет на присоединение кислорода к другому гему той же молекулы. Это явление было известно задолго до работ Перутца и установления структуры гемоглобина и механизма его реакции с кислородом. Оно получило название гем-гем взаимодействия. Физиологический смысл гем-гем взаимодействия очевиден. Сигмоидная форма кривой диссоциации создает условия максимальной отдачи кислорода при переносе гемоглобина от легких с высоким значением pO2 к тканям с низким значением pO2. Для человека значения pO2 артериальной и венозной крови в нормальных условиях (Т 37°С, pH 7,4) равны соответственно 100 и 40 ммHg. При этом (рис.2, б) гемоглобин отдает тканям 23% связанного кислорода (степень оксигенации меняется от 98 до 75%). При отсутствии гем-гем взаимодействия для одногемового миоглобина (рис.2, а) эта величина не превышает 5%. Миоглобин поэтому служит не переносчиком, а депо кислорода и отдает его мышечной ткани лишь при резкой гипоксии, когда насыщение ткани кислородом падает до недопустимо низкого значения. [2]
Логарифмическая анаморфоза кривой диссоциации гемоглобина человека представлена на рис.3, б. В этом случае начало кривой представляет собой прямую под углом 45° к координатным осям, как и для миоглобина: первые молекулы кислорода соединяются в основном с молекулами гемоглобина, еще не содержащими кислорода, и гемы таким образом оксигенируются независимо. Это свидетельствуют о том, что гем-гем взаимодействие обусловлено не просто наличием нескольких гемов в молекуле, а тем, что после оксигенации одного гема меняются условия оксигенации других гемов той же молекулы. Затем наклон кривой увеличивается. Тангенс угла максимального наклона получил название коэффициента Хилла (n), который отражает степень кооперативности процесса. Для миоглобина n=1, а для гемоглобина человека (в норме) n~3. Вблизи области полного насыщения гемоглобина кислородом наклон кривой снова становится равным 45° (большинство молекул гемоглобина либо не содержат свободных гемов, либо имеют лишь один гем, способный присоединить кислород).
Реферат опубликован: 8/04/2005 (18398 прочтено)