Страница: 1/6
Содержание
- Введение
- Химическая структура цефалоспоринов
- Механизм действия цефалоспоринов
- Механизмы формирования резистентности к цефалоспоринам
- Классификация цефалоспоринов
- Фармакология цефалоспоринов
- Цефалоспорины 1-го поколения
- Цефалоспорины 2-го поколения
- Цефалоспорины 3-го поколения
- Цефалоспорины 4-го поколения
- Побочные эффекты цефалоспоринов
- Список используемой литературы
Введение
Создание и внедрение в клиническую практику антибиотиков класса цефалоспоринов явилось, безусловно, одним из важнейших событием в истории химиотерапии бактериальных инфекции. В настоящее время, очевидно, что по ряду важнейших параметров: спектру антибактериального действия, фармакокинетике, безопасности и др., цефалоспорины превосходят антибиотики многих других классов, чем и объясняется их статус антибактериальных средств, наиболее широко применяемых во многих странах мира.
Рождение цефалоспоринов справедливо связывают с работами G.Brotzu, предположившего в начале 40-х годов, что периодическое самоочищение сточных вод в Сардинии обусловлено ингибирующей активностью особых микроорганизмов. В 1945 г. ему удалось выделить гриб Cephalosporium acremonium (в настоящее время именуемый Acremonium chrysogenum), обладавший выраженной антибактериальной активностью по отношению к грамположительным и грамотрицательным микроорганизмам. Следующим этапом (с 1955 по 1962 г.) явилось детальное изучение С.acremonium рабочими группами H.Florey и E.P.Abraham. Из продуктов обмена веществ этого гриба удалось выделить бактерицидную субстанцию - цефалоспорин С, ставшую исходным веществом для получения 7-аминоцефалоспорановой кислоты - структурной основы цефалоспоринов.
В 1962 г. в клиническую практику был введен первый антибиотик класса цефалоспоринов - цефалоридин, однако широкое клиническое применение цефалоспоринов началось только в конце 70-х годов. В настоящее время насчитывается более 50 цефалоспоринов (рис. 1), поэтому нет ничего удивительного в том, что врач испытывает немалые затруднения при выборе нужного препарата в соответствии с показаниями. В связи с этим целью настоящей лекции является объективная оценка возможностей цефалоспоринов в современной клинической практике.

Рис.1. "Древо" цефалоспоринов (по Periti, 1996)
Химическая структура цефалоспоринов
Цефалоспорины представляют собой бициклические соединения, состоящие из бета-лактамного и дигидротиазинового колец. Оба кольца и составляют 7-аминоцефалоспорановую кислоту (7-АЦК) - общее ядро молекулы цефалоспоринов. При этом модификация химической структуры 7-АЦК сопровождается существенными изменениями свойств (антибактериальной активности, параметров фармакокинетики и пр.) соответствующего соединения (рис. 2).
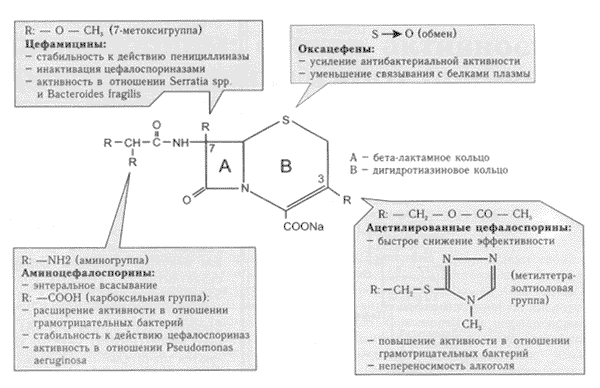 Рис.2.
Цефалоспорины: связь между химической структурой и эффектом
Рис.2.
Цефалоспорины: связь между химической структурой и эффектом
(по W.Graninger, 1994)
Механизм действия цефалоспоринов
Антибактериальная активность цефалоспоринов, как и других бета-лактамных антибиотиков, по крайней мере частично, обусловлена торможением синтеза пептидогликана - структурной основы микробной стенки. Пептидогликаны представляют собой длинные полисахаридные цепи со своеобразной сетчатой пространственной конформацией, в которых чередуются остатки N-ацетилглюкозамина (NAG) и N-ацетилмураминовой кислоты (NAM). NAG- и NAM-пентапептидные остатки пептидогликанов синтезируются в цитоплазме микробной клетки и транспортируются через цитоплазматическую мембрану. Далее эти остатки встраиваются в существующую пептидогликанную сеть (в процессе роста и деления клетки) с участием различных энзимов - транспептидаз, карбоксипептидаз, эндопептидаз. Собственно эти энзимы, находящиеся в цитоплазматической мембране, являются местом реализации антибактериальной активности (мишенями) бета-лактамных антибиотиков, в том числе цефалоспоринов; они получили название "пенициллинсвязывающие белки" (penicillin-binding proteins - РВР). В результате образования "длительной" ковалентной связи бета-лактамного антибиотика и РВР последние инактивируются. При этом эффект назначаемого бета-лактамного антибиотика зависит от того, какие РВР инактивируются и какую роль они играют в синтезе пептидогликана и выживании микробной клетки. Важно также подчеркнуть, что бактерицидный эффект цефалоспоринов реализуется только в процессе роста и размножения микроорганизмов, тогда как "покоящиеся" клетки неуязвимы для действия антибиотиков.
Механизмы формирования резистентности к цефалоспоринам
Устойчивость микроорганизмов к действию цефалоспоринов может быть связана с одним из следующих механизмов: а) видоизменением (модификация) РВР со снижением аффинности (сродства) к ним цефалоспоринов; б) гидролизной инактивацией антибиотика (бета-лактамазами); в) нарушением проницаемости внешних структур микробной клетки для антибиотика и затруднением его связывания с "мишенью" - РВР.
У грамположительных микроорганизмов цитоплазматическая мембрана относительно порозна и непосредственно прилежит к пептидогликанному матриксу, с связи с чем цефалоспорины достаточно легко достигают РВР. В противоположность этому, наружная мембрана грамотрицательных микроорганизмов имеет существенно боле% сложную "конструкцию": состоит из липидов, полисахаридов и белков, что является препятствием для проникновения цефалоспоринов в периплазматическое пространство микробной клетки (рис. 3). Цефалоспорины "проходят" сквозь наружную мембрану микробной клетки через так называемые пориновые каналы. В связи с этим, очевидно, уменьшение проницаемости пориновых каналов может привести к формированию антибиотикорезистентности.
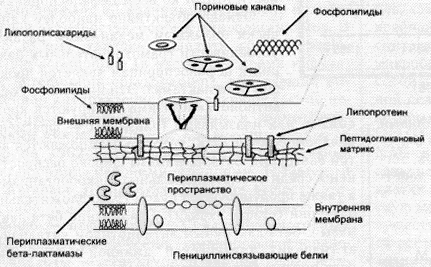
Рис.3. Схема строения бактериальной стенки грамотрицательных микроорганизмов
(по P.A.James, 1996)
Снижение сродства РВР к бета-лактамным антибиотикам рассматривают как ведущий механизм формирования резистентности Neisseria gonorrhea и Streptococcus pneumoniae к пенициллину. Метициллинрезистентные штаммы Staphylococcus aureus (MRSA) продуцируют РВР2 (РВР2а), которые характеризуются значительным снижением аффинности к пенициллиназарезистентным пенициллинам и цефалоспоринам. Способность этих "новых" РВР2а к замещению эссенциальных РВР (с более высоким сродством к бета-лактамам) в конце концов, приводит к формированию устойчивости MRSA ко всем цефалоспоринам.
Однако объективно наиболее клинически значимым механизмом развития устойчивости грамотрицательных бактерий к цефалоспоринам является продукция бета-лактамаз. Эти инактивирующие антибиотики энзимы кодируются хромосомами или плазмидами (плазмиды - фрагменты внехромосомной ДНК, которые размножаются внутри бактерий). Бета-лактамазы широко распространены среди грамотрицательных микроорганизмов, а также продуцируются рядом грамположительных бактерий (стафилококки). Связывание бета-лактамазы с бета-лактамным антибиотиком катализирует гидролиз "критической" аминной связи лактамного кольца, что и приводит к инактивации антибиотика.
Грамположительные микроорганизмы высвобождают бета-лактамазы непосредственно в окружающее их внеклеточное пространство. При этом известно, что большинство цефалоспоринов (за исключением, пожалуй, цефалоридина) достаточно устойчиво к гидролизующему действию стафилококковой бета-лактамазы. В связи с этим антистафилококковая активность цефалоспоринов зависит главным образом от их сродства к эссенциальным стафилококковым РВР. Так, например, цефамицины и цефтазидим, являясь достаточно бета-лактамазостабильными, демонстрируют низкую антистафилококковую активность вследствие низкого сродства к РВР S.aureus.
Бета-лактамазная резистентность грамотрицательных бактерий к цефалоспоринам имеет более сложный характер. У этих микроорганизмов бета-лактамазы "заключены" в периплазматическом пространстве. Важно подчеркнуть, что повышение продукции бета-лактамаз или новообразование энзимов с повышенным сродством к антибиотику является основной причиной распространения цефалоспоринрезистентных штаммов среди грамотрицательных микроорганизмов. При этом высокий уровень продукции TEM-I или SHV-I*(Чаще всего в названиях бета-лактамаз учитываются основные субстраты-антибиотики, фамилии исследователей или пациентов. Так, например, термин "ТЕМ" стали использовать для обозначения плазмидных энзимов, первоначально выделенных из микроорганизмов у больного Temorina, a SHV - англоязычная аббревиатура термина "сульфгидрильная вариабельность”), двух наиболее часто встречающихся плазмидассоцируемых бета-лактамаз бактерий семейства Enterobacteriaceae (Escherichia coli, Salmonella spp., Shigella spp., Klebsiella spp., Enterobacter spp. и др.), ассоциируется с формированием резистентности не только к пенициллинам - ингибиторам бета-лактамаз, но и к цефалотину, цефамандолу, цефоперазону.
Отдельные представители семейства Enterobacteriaceae (Enterobacter spp., Citrobacter freundii, Morganella morganii, Serratia marcescens, Providencia spp.), а также Pseudomonas aeruginosa демонстрируют способность к продукции "индуцибельных" хромосомных цефалоспориназ, характеризующихся высоким сродством к цефамицинам и цефалоспоринам 3-го поколения. Индукция или стабильное "дерепрессирование" этих хромосомных бета-лактамаз в период "давления" (применения) цефамицинов или цефалоспоринов 3-го поколения в итоге приведет к формированию резистентности ко всем доступным цефалоспоринам. Распространение данной формы резистентности увеличивается в случаях лечения инфекций, прежде всего вызываемых Enterobacter cloaceae и Pseudomonas aeruginosa, цефалоспоринами широкого спектра действия.
Реферат опубликован: 16/06/2005 (15383 прочтено)