Страница: 1/2
Введение
Гемоглобин имеет важнейшее биологическое значение как наиболее широко распространенный среди современных животных дыхательный пигмент, обладающий уникальными свойствами, связанными с транспортом кислорода и углекислоты.
Одна из важнейших функций организма, определяющая характер и уровень энергетических процессов,— дыхание или обеспечение его кислородом. Осуществляется эта функция, как известно, неодинаково у различных групп животных. Малоактивные животные (простейшие, губки, кишечнополостные, некоторые черви, моллюски, ракообразные) обеспечиваются кислородом путем простой диффузии через поверхность тела. Высокоактивные животные, нуждающиеся в больших количествах кислорода, обладают соответствующими эффективными механизмами, обеспечивающими поступление кислорода в организм. В частности, насекомые и некоторые другие представители членистоногих обладают трахейной системой, пронизывающей все тело и доставляющей кислород непосредственно тканям и клеткам самых отдаленных от поверхности частей тела. Это чрезвычайно эффективный способ, обеспечивающий высокий уровень энергетических процессов в организме этой группы животных.
Принципиально отличен от первых двух способ обеспечения организма кислородом при помощи дыхательных пигментов крови (позвоночные животные, некоторые черви) или гемолимфы (часть червей, ракообразных, моллюсков). В то время как при дыхании через поверхность тела и при помощи трахей кислород доставляется непосредственно клеткам тела, у пигментсодержащих животных кислород из органов аэрации доставляется клеткам тела посредством дыхательных пигментов крови или гемолимфы.
Следовательно, помимо органов дыхания, в которых происходит обмен газов, у этой группы животных кровь содержит особые вещества — дыхательные пигменты, способные связывать кислород. Кроме того, у них имеется сосудистая система, по которой движется кровь, вступая в контакт со всеми частями тела.
В настоящее время известно четыре дыхательных пигмента, встречающихся в организме современных животных: гемоглобин, гемоцианин, хлорокруорин и гемэритрин. Из них только гемоцианин содержит медь, в состав остальных входит железо. Близко родственны по структуре гемоглобин и хлорокруорин.
Гемоглобин является основным дыхательным пигментом, распространенным среди всех групп животных
Структура гемоглобина
В состав молекулы гемоглобина входят четыре одинаковые гемовые группы. Гем представляет собой протопорфирин, содержащий центрально расположенный ион двухвалентного железа. Молекула протопорфирина состоит из четырех пиррольных колец, связанных метиновыми мостиками; к кольцам присоединены боковые цепи характерного строения. Ключевую роль в активности гемоглобина играет ион железа, расположенный в центре молекулы протопорфирина. Соединение с этим ионом посредством двух координационных связей и двух связей, образовавшихся вследствие замещения водорода, превращает протопорфирин в гем. Структура гема целиком расположена в одной плоскости. В процессе переноса кислорода гемоглобином молекула Од обратимо связывается с гемом, при этом валентность железа не изменяется.
Присоединяя кислород, гемоглобин (Hb) превращается в оксигемоглобин (HbOx). Чтобы подчеркнуть тот факт, что валентность железа при этом связывании не меняется, реакцию называют не окислением, а оксигенацией; обратный процесс называется дезоксигенацией. Когда хотят специально отметить, что гемоглобин не связан с кислородом, его называют дезоксигемоглобином.
Гем может подвергаться не только оксигенации, но и истинному окислению. При этом железо переходит из двухвалентного состояния в трехвалентное. Окисленный гем называется гематином (мет-гемом), а вся полипептидная молекула в целом – метгемоглобином. В крови человека метгемоглобин содержится в незначительных количествах, но при некоторых заболеваниях и отравлениях определенными ядами его содержание возрастает. Опасность таких состояний заключается в том, что окисленный гемоглобин не способен переносить кислород к тканям.
Видовые свойства гемоглобинов крови различных животных (различные формы кристаллов, сродство гемоглобина к кислороду и др.) определяются особенностями глобина, так как гемы всех исследованных гемоглобинов оказались идентичными.
Гемоглобин миноги
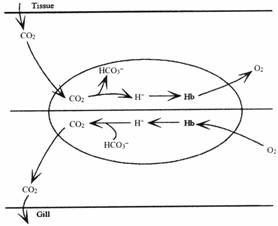
Функционирование гемоглобина в эритроцитах миноги предлагает уникальное среди позвоночных решение газового транспорта. Гемоглобин миноги в неповрежденных эритроцитах находится в олигомерном\мономерном равновесии и обладает сходным с остальными активными рыбами кислородным сродством. Проницаемость мембраны эритроцитов миноги для кислотных и основных эквивалентов очень низка и таким образом бикарбонат плазмы не может в достаточной степени дегидрироваться до CO2 пока кровь находится в жабрах. Это потенциальное ограничение на выведение CO2 преодолевается высоким внутриэритроцитарным pH и изменениями pH внутри эритроцита связанными с оксигенацией из-за относительной непроницаемости мембраны эритроцитов для кислотных эквивалентов внутриэритроцитарный гемоглобин не принимает участие в кислотно-основном забуферивании внеклеточного компартмента. Вследствие этого внеклеточные кислотные заряды вызывают заметные изменения pH плазмы.
Функции гемоглобина миног в основном изучались в разбавленных растворах. На основании этих исследований ясно, что гемоглобин миноги в растворах показывает как кооперативность, так и эффекты Бора и Хэлдена. Кроме того, сродство гемоглобина к кислороду у миноги зависти от концентрации гемоглобина в растворе. Это открытие, а так же наблюдение, что молекулярная масса деоксигенированного гемоглобина больше, чем оксигенированного показывают, что в миногах кооперативность связывания кислорода и влияние протонов на сродство гемоглобина к кислороду вызваны обратимыми реакциями ассоциации\диссоциации олигомеров\мономеров. Это явление противоположно ситуации с гемоглобином у всех других позвоночных, у которых эти эффекты являются результатом конформационных изменений тетрамерной молекулы.
Поведение гемоглобина миноги в растворе может быть объяснено следующей моделью: димерная\олигомерная форма молекулы имеет низкое сродство к кислороду вследствие ограничений, следующих из взаимодействий между субъединицами. В димере связывание кислорода ослабляет эти взаимодействия, молекула диссоциирует на субъединицы, вследствие чего сродство к кислороду каждой субъединицы возрастает. Вероятно, что дистальный гистидин (Е7) вовлечен в аллостерический переход: в мономерной форме он находится во внутренней позиции внутри гемового кармана (где он может образовывать водородную связь с кислородом), а в димерной форме гистидин (Е7) находится в наружном положении (где он формирует водородную связь с аспартатом и глутаматом соседней молекулы). Эффект Бора в таком случае объясняется различием между значениями pK гистидина во внутреннем положении (5,5-6,0) и во внешнем (ок. 8,0).
До недавнего времени было затруднительно интерпретировать данные по связыванию кислорода цельной кровью на основе поведения гемоглобина в растворе: например, сродство крови миноги к кислороду оказалось намного больше, чем ожидаемое на основании поведения гемоглобина в растворе, а так же численное значение фактора Бора, оцененное для всей крови оказалось намного ниже, ожидаемого на основании данных из раствора.
Противоречие между данными по связыванию кислорода гемоглобином в растворах и в эритроцитах было разрешено после открытия уникального среди позвоночных свойства мембраны эритроцитов миноги: ее проницаемость для кислотных и основных эквивалентов, а так же для хлорида очень низка. Вследствие очень незначительного пассивного транспорта кислотных эквивалентов через эритроцитарную мембрану внутриклеточные значения pH поддерживаются на высоком уровне в основном Na+/H+ обменом через мембрану.
Это уникальное свойство мембраны влияет на функционирование гемоглобина в эритроцитах миноги, на газовый транспорт и кислотно-основную регуляцию.
Сродство гемоглобина к кислороду у миноги
Сродство гемоглобина к кислороду у миноги такое же, как и у других активных рыб. Поддержание этого сродства к кислороду требует, чтобы внутриэритроцинарный pH сохранялся на более высоком уровне, чем у костных рыб. Lampetra fluviatilis и Petromyzon marinus поддерживают внутриэритроцитарный pH используя механизм, подобный натриево-протоновому обмену.
Кооперативность связывания кислорода
Деоксигенированный гемоглобин миноги находится в олигомерной форме, обладающей низким сродством к кислороду, которая стабилизирована при низких значениях pH (при очень низком pH диссоциация не наблюдается). При увеличении значений pH взаимодействия между индивидуальными цепочками олигомеров ослабляются, и оксигенация вызывает диссоциацию обладающих низким сродством к кислороду олигомеров на обладающие высоким сродством мономеры. Чем выше значение pH, тем ниже требуемое для диссоциации олигомеров парциальное давление CO2.
Эффект протонов на функции гемоглобина
 Ранние исследования влияния pH на кислород-связывающие свойства крови миноги
допускали, что фактор Бора мал (примерно от -0,1 до -0,3). Однако в тех исследованиях
сродство гемоглобина к кислороду рассматривалось как функция от внеклеточного
pH, поскольку в то время уникальность регуляции внутриклеточного pH в эритроцитах
миноги не была известна. Более поздние исследования, в которых изменение сродства
к кислороду соотносили с внутриклеточным pH показали, что фактор Бора достаточно
велик. У Petromyzon marinus при 10° С и pHi=7.3-7.6 значение фактора Бора (DlogP50/DpHi)
оказались равны –0,63, а в Lampetra fluviatilis при 18°С и при pHi=6,7-8,0 фактор
Бора равен 1,03. Для Lampetra fluviatilis это значение показывает освобождение
одного протона на один мономерный гемоглобин. Подобное же значение было получено
на основе небикарбонатной буферной способности эритроцита, внутриэритроцитарной
концентрации гемоглобина и внутриклеточных изменений pH при оксигенации. Таким
образом эффект Бора гемоглобина миноги происходит из-за выпуска одного протона
на одну цепочку гемоглобина при оксигенации.
Ранние исследования влияния pH на кислород-связывающие свойства крови миноги
допускали, что фактор Бора мал (примерно от -0,1 до -0,3). Однако в тех исследованиях
сродство гемоглобина к кислороду рассматривалось как функция от внеклеточного
pH, поскольку в то время уникальность регуляции внутриклеточного pH в эритроцитах
миноги не была известна. Более поздние исследования, в которых изменение сродства
к кислороду соотносили с внутриклеточным pH показали, что фактор Бора достаточно
велик. У Petromyzon marinus при 10° С и pHi=7.3-7.6 значение фактора Бора (DlogP50/DpHi)
оказались равны –0,63, а в Lampetra fluviatilis при 18°С и при pHi=6,7-8,0 фактор
Бора равен 1,03. Для Lampetra fluviatilis это значение показывает освобождение
одного протона на один мономерный гемоглобин. Подобное же значение было получено
на основе небикарбонатной буферной способности эритроцита, внутриэритроцитарной
концентрации гемоглобина и внутриклеточных изменений pH при оксигенации. Таким
образом эффект Бора гемоглобина миноги происходит из-за выпуска одного протона
на одну цепочку гемоглобина при оксигенации.
Реферат опубликован: 16/06/2005 (5795 прочтено)