Страница: 2/3
Рис. 1 Линии перерезок ствола мозга
Первый препарат был назван Бремером cerveau isolee, а второй encephale isolee (изолированный передний мозг и изолированный головной мозг). Межколликулярная перерезка оставляет ниже места разреза почти всю ретикулярную формацию; перерезка же ниже продолговатого мозга оставляет в связи с высшими отделами мозга все стволовые структуры. У животного с ceryeau isolee остаются связанными с корой головного мозга такие важнейшие афферентные системы, как обонятельная и зрительная. Однако такое животное не реагирует на световые и обонятельные раздражения; глаза у него закрыты, и оно фактически не вступает в контакт с внешним миром. У животного же с encephale isolee реакции полностью сохраняются; такое животное открывает глаза на свет, реагирует на запахи, т.е. ведет себя как бодрствующее, а не как спящее животное.
|
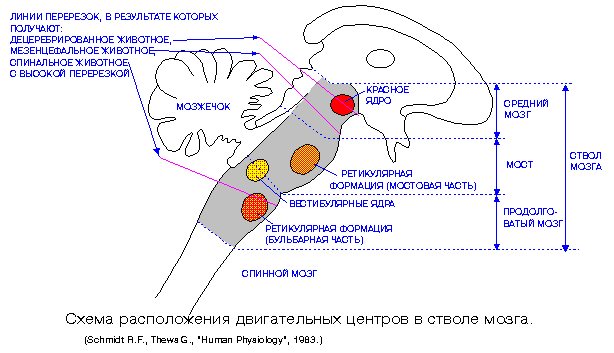
Однако дальнейшие исследования показали, что для поддержания бодрствующего состояния коры больших полушарий важно не просто поступление к ней импульсации по афферентным системам. Если сделать перерезку ствола мозга так, чтобы не повредить основные афферентные системы (например, систему медиальной петли), но перерезать восходящие связи ретикулярной формации, то животное тем не менее впадает в сонное состояние, конечный мозг перестает активно функционировать.
Следовательно, для поддержания бодрствующего состояния конечного мозга важно, чтобы афферентная импульсация первоначально активировала ретикулярные структуры мозгового ствола. Влияния же из ретикулярных структур по восходящим путям каким то образом определяют функциональное состояние конечного мозга. Проверить такой вывод можно путем прямого раздражения ретикулярных структур. Такое раздражение через погружные электроды было проведено Дж.Моруцци и X.Мегоуном и затем воспроизводилось во многих лабораториях в условиях хронического или полухронического опыта. Оно всегда дает однозначные результаты в виде характерной поведенческой реакции животного. Если животное находится в сонном состоянии оно просыпается, у него появляется ориентировочная реакция. После прекращения раздражения животное снова возвращается в сонное состояние. Переход от сонного к бодрствующему состоянию в период раздражения ретикулярных структур отчетливо проявляется не только в поведенческих реакциях, он может быть зарегистрирован по объективным критериям деятельности коры больших полушарий, в первую очередь по изменениям ее электрической активности.
Для коры больших полушарий характерна постоянная электрическая активность (ее запись называется электрокортикограммой). Эта электрическая активность состоит из небольших по амплитуде (30-100 мкВ) колебаний, которые легко отводятся не только от открытой поверхности мозга, но и от кожи головы. У человека в спокойном дремотном состоянии такие колебания имеют частоту 8-10 в секунду и являются довольно регулярными (альфа-ритм). У высших позвоночных этот ритм менее регулярен, а частота колебаний варьирует от 6-8 у кролика до 15--20 у собаки и обезьяны. Во время активности регулярные колебания сменяются сразу же значительно меньшими по амплитуде и более высокочастотными колебаниями (бета-ритм). Появление периодических больших колебаний ясно говорит о том, что электрическая активность каких-то элементов в коре развивается синхронно. Когда правильные колебания большой амплитуды сменяются низковольтными, частыми колебаниями, то это, очевидно, говорит о том, что клеточные элементы коры начинают функционировать менее синхронно, поэтому такой тип активности называется реакцией десинхронизации. Таким образом, переход от спокойного, неактивного состояния коры к активному связан в электрическом отношении с переходом от синхронизированной активности ее клеток к десинхронизированной.
Характерным эффектом восходящих ретикулярных влияний на корковую электрическую активность является именно реакция десинхронизации. Эта реакция закономерно сопровождает описанную выше поведенческую реакцию, характерную для ретикулярных влияний. Реакция десинхронизации не ограничивается каким-нибудь одним участком коры, а регистрируется от больших ее областей. Это говорит о том, что восходящие ретикулярные влияния являются генерализованными.
Описанные изменения в электроэнцефалограмме не являются единственным электрическим проявлением восходящих ретикулярных влияний. В определенных условиях можно выявить более прямые эффекты ретикулярной импульсации, поступающей к коре больших полушарий. Они впервые были описаны в 1940 г. американскими исследователями А.Форбсом и Б.Мориссоном, которые исследовали вызванную электрическую активность коры при различных афферентных влияниях. При раздражении какой-либо афферентной системы в соответствующей проекционной зоне коры обнаруживается электрический ответ, указывающий на приход к этой области афферентной волны, этот ответ получил название первичного ответа. Кроме этого локального ответа, афферентное раздражение вызывает длиннолатентный ответ, возникающий в обширных областях коры мозга. Этот ответ Форбс и Мориссон назвали вторичным ответом.
То обстоятельство, что вторичные ответы возникают со скрытым периодом, значительно превышающим скрытый период первичного ответа, ясно говорит о том, что они связаны с поступлением афферентной волны в кору не по прямым, а по каким-то окольным связям, через дополнительные синаптические переключения. Позже, когда было применено прямое раздражение ретикулярной формации, было показано, что оно может вызвать ответ такого же типа. Это позволяет заключить, что вторичный ответ является электрическим проявлением поступления в кору больших полушарий афферентное волны через ретикулокортикальные связи.
Через ствол мозга проходят прямые афферентные пути, которые после синаптического перерыва в таламусе поступают в кору больших полушарий. Приходящая по ним афферентная волна вызывает первичный электрический ответ с соответствующей проекционной зоне с коротким скрытым периодом. Одновременно афферентная волна по коллатералям ответвляется в ретикулярную формацию и активирует ее нейроны. Затем по восходящим путям от нейронов ретикулярной формации импульсация тоже поступает в кору, но уже в виде задержанной реакции, возникающей с большим скрытым периодом. Эта реакция охватывает не только проекционную зону, но и большие участки коры, вызывая в них какие-то изменения, важные для бодрствующего состояния.
Нисходящие функции ретикулярной формации включают в себя, как правило, облегчающие и тормозящие компоненты, которые осуществляются по реципрокному принципу. Швейцарский физиолог В. Гесс (1929) впервые показал, что в стволе мозга можно найти точки, при раздражении которых у животного наступает сон. Гесс назвал эти точки центрами сна. Позже Дж.Моруцци и сотр. (1941) также обнаружили, что, раздражая некоторые участки ретикулярной формации заднего мозга, можно у животных вызвать в коре вместо десинхронизации синхронизацию электрических колебаний н соответственно перевести животное из бодрствующего состояния в пассивное, сонное. Поэтому можно думать, что в составе восходящих путей ретикулярной формации действительно существуют не только активирующие, но и инактивирующие подразделения, последние каким-то образом снижают возбудимость нейронов конечного мозга.
Нейронная организация восходящей системы ретикулярной формации не вполне ясна. При разрушении ретикулярных структур среднего и заднего мозга в коре больших полушарий не обнаруживается существенного количества дегенерирующих окончаний, которые могли бы быть отнесены к прямым ретикулярным волокнам. Значительная дегенерация окончаний в коре возникает только при разрушении неспецифических ядер таламуса. Поэтому возможно, что восходящие ретикулярные влияния передаются в кору больших полушарий не по прямым путям, а через какие-то промежуточные синаптические соединения, локализованные, вероятно, в промежуточном мозге.
Любопытно отметить, что гистологические и электрофизиологические данные указывают на характерную деталь хода аксонов многих ретикулярных нейронов. Аксоны нейронов гигантоклеточного ядра, т. е. основного ядра ретикулярной формации, очень часто делятся Т-образно, и один из отростков идет вниз, образуя ретикулоспинальный путь, а второй вверх, направляясь в верхние отделы головного мозга. Создается такое впечатление, что и восходящие, и нисходящие функции ретикулярной формации могут быть связаны с деятельностью одних и тех же нейронов. По функциональным свойствам ретикулярные структуры, создающие восходящие влияния, также имеют много общего со структурами, обеспечивающими нисходящие влияния. Восходящие влияния несомненно носят тонический характер, они легко изменяются гуморальными факторами и высокочувствительны к фармакологическим веществам. Снотворное и наркотическое действие барбитуратов основано, по-видимому, именно на блокировании в первую очередь восходящих влияний ретикулярной формации.
4. Свойства ретикулярных нейронов
Большой интерес представляют результаты исследований общих закономерностей
деятельности нейронов ретикулярной формации. Этим исследованиям положили начало
работы Дж.Моруцци, который впервые подробно описал функциональные свойства ретикулярных
нейронов, использовав при этом внеклеточное микроэлектродное отведение. При
этом сразу же обратила на себя внимание их способность к стойкой импульсной
активности. Если деятельность ретикулярной формации исследуется в условиях отсутствия
наркоза, то большинство ее нейронов постоянно генерирует нервные разряды с частотой
порядка 5-10 в секунду. Различные афферентные влияния суммируются с этой фоновой
активностью ретикулярных нейронов, вызывая в одних из них ее учащение, а в других
торможение. Причина постоянной фоновой активности в этих условиях, по-видимому,
двоякая. С одной стороны, она может быть связана с высокой химической чувствительностью
мембраны ретикулярных клеток и постоянной ее деполяризацией гуморальными химическими
факторами. С другой стороны, она может определяться особенностями афферентных
связей ретикулярных нейронов, а именно, конвергенцией к ним коллатералей от
огромного количества различных сенсорных путей,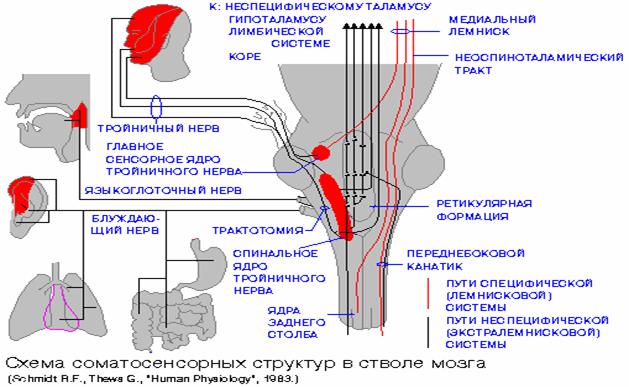
поэтому даже в случае, когда организм не подвергается действию каких-нибудь
специальных раздражителей в ретикулярную формацию могут непрерывно поступать
импульсы от всевозможных неконтролируемых воздействий (Рис. 2). Эти импульсы,
приходя к клеткам через многочисленные синаптические входы, вызывают также дополнительную
деполяризацию мембраны. В связи с таким характером активности ретикулярных клеток
их влияния на другие структуры также носят постоянный, тонический характер.
Если, например, искусственно прервать связь ретикулярной формации со спинным
мозгом, то это сразу же вызовет существенные стойкие изменения рефлекторной
деятельности последнего; в частности, облегчаются рефлексы, осуществляющиеся
по полисинаптическим путям. Это ясно говорит о том, что спинальные нервные элементы
находятся под тоническим, преимущественно тормозящим. контролем со стороны ретикулярной
формации.
|
Реферат опубликован: 15/06/2005 (13411 прочтено)