Страница: 4/5
В качестве прокариотических векторов используются плазмиды, бактериофаги; в качестве эукариотических векторов применяют вирусы животных и растений, векторы на основе 2 мкм дрожжей и митохондрий и ряд искусственно сконструированных векторов, способных реплицироваться как в бактериальных, так и в эукариотических клетках (челночные векторы).
Плазмиды - это внехромосомные генетические элементы про- и эукариот, которые автономно реплицируются в клетках. Большинство плазмидных векторов получено на основе природных плазмид ColE1, pMB1 и p15A.
Бактериальные плазмиды делят на два класса. Одни плазмиды (например, хорошо изученный фактор F, определяющий пол у E.coli) сами способны переходить из клетки в клетку, другие такой способностью не обладают. По ряду причин, и прежде всего для предотвращения неконтролируемого распространения потенциально опасного генетического материала, подавляющее большинство бактериальных плазмидных векторов создано на базе плазмид второго класса. Многие природные плазмиды уже содержат гены, определяющие устойчивость клеток к антибиотикам (продукты этих генов - ферменты, модифицирующие или расщепляющие антибиотические вещества). Кроме того, в эти плазмиды при конструировании векторов вводятся дополнительные гены, определяющие устойчивость к другим антибиотикам.
На рис. 4 показан один из самых распространенных плазмидных векторов E.coli - pBR322. Он сконструирован на базе детально изученной плазмиды E.coli - колициногенного фактора ColE1 - и содержит ориджин репликации этой плазмиды. Особенность плазмиды ColE1 (и pBR322 соответственно) состоит в том, что в присутствии ингибитора синтеза белка антибиотика хлорамфеникола (опосредованно ингибирующего репликацию хозяйской хромосомы) ее число в E.coli возрастает от 20-50 до 1000 молекул на клетку, что позволяет получать большие количества клонируемого гена. При конструировании вектора pBR322 из исходных плазмид был делегирован целый ряд "лишних" сайтов для рестриктаз.
В настоящее время наряду с множеством удобных векторных систем для E.coli сконструированы плазмидные векторы для ряда других грамотрицательных бактерий (в том числе таких промышленно важных, как Pseudomonas, Rhizobium и Azotobacter), грамположительных бактерий (Bacillus), низших грибов (дрожжи) и растений.
Плазмидные векторы удобны для клонирования относительно небольших фрагментов (до 10 тыс. пар оснований) геномов небольших размеров. Если же требуется получить клонотеку (или библиотеку) генов высших растений и животных, общая длина генома которых достигает огромных размеров, то обычные плазмидные векторы для этих целей непригодны. Проблему создания библиотек генов для высших эукариот удалось решить с использованием в качестве клонирующих векторов производных бактериофага l.
Среди фаговых векторов наиболее удобные системы были созданы на базе геномов бактериофагов l и М13 E.coli. ДНК этих фагов содержит протяженные области, которые можно делегировать или заменить на чужеродную ДНК, не затрагивая их способности реплицироваться в клетках E.coli. При конструировании семейства векторов на базе ДНК l фага из нее сначала (путем делений коротких участков ДНК) были удалены многие сайты рестрикции из области, не существенной для репликации ДНК, и оставлены такие сайты в области, предназначенной для встраивания чужеродной ДНК. В эту же область часто встраивают маркерные гены, позволяющие отличить рекомбинантную ДНК от исходного вектора. Такие векторы широко используются для получения "библиотек генов". Размеры замещаемого фрагмента фаговой ДНК и соответственно встраиваемого участка чужеродной ДНК ограничены 15-17 тыс. нуклеотидных остатков, так как рекомбинантный фаго -
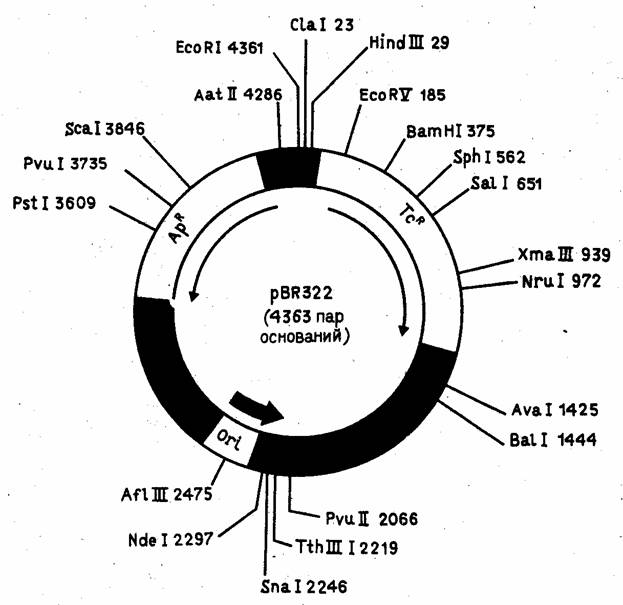
Рисунок 4. Детальная рестрикционная карта плазмиды pBR322.
вый геном, который на 10% больше или на 75% меньше генома дикого l фага, уже не может быть упакован в фаговые частицы.
Таких ограничений теоретически не существует для векторов, сконструированных на базе нитчатого бактериофага М13. Описаны случаи, когда в геном этого фага была встроена чужеродная ДНК длиной около 40 тыс. нуклеотидных остатков. Известно, однако, что фаг М13 становится нестабильным, когда длина чужеродной ДНК превышает 5 тыс. нуклеотидных остатков. Фактически же векторы, полученные из ДНК фага М13, используются главным образом для секвенирования и мутагенеза генов, и размеры встраиваемых в них фрагментов намного меньше.
Эти векторы конструируются из реплекативной (двутяжевой) формы ДНК фага М13, в которую встроены "полилинкерные" участки (пример такой конструкции показан на рис. 5). В фаговую частицу ДНК включается в виде однотяжевой молекулы. Таким образом, этот вектор позволяет получать клонированный ген или его фрагмент как в двутяжевой, так и в однотяжевой форме. Однотяжевые формы рекомбинантных ДНК широко используются в настоящее время при определении нуклеотидной последовательности ДНК методом Сэнгера и для олигодезоксинуклеотид-направленного мутагенеза генов.
Перенос чужеродных генов в клетки животных осуществляется с помощью векторов, полученных из ДНК ряда хорошо изученных вирусов животных - SV40, некоторых аденовирусов, вируса папиломы быка, вируса оспы и так далее. Конструирование этих векторов проводится по стандартной схеме: удаление "лишних" сайтов для рестриктаз, введение маркерных генов в области ДНК, не существенные для ее репликации (например, гена тимидин-киназы (tk) из HSV (вируса герпеса)), введение регуляторных районов, повышающих уровень экспрессии генов.
Удобными оказались так называемые "челночные векторы", способные реплицироваться как в клетках животных, так и в клетках бактерий. Их получают, сшивая друг с другом большие сегменты векторов животных и бактерий (например, SV40 и pBR322) так, чтобы районы, ответственные за репликацию ДНК, остались незатронутыми. Это позволяет проводить основные операции по конструированию вектора в бактериальной клетке (что технически намного проще), а затем полученную рекомбинантную ДНК использовать для клонирования генов в животной клетке.
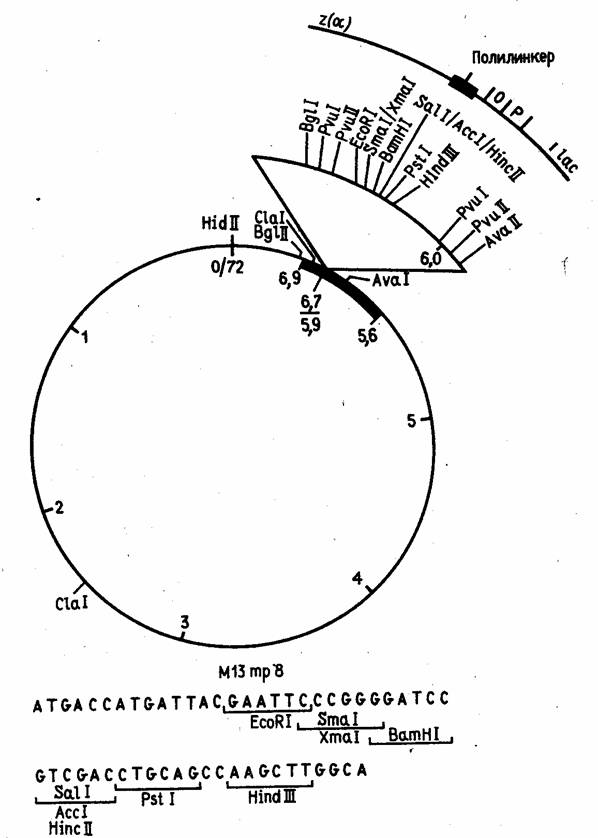
Рисунок 5. Рестрикционная карта вектора М13 mp8.
1.3. ВЫБОР СИСТЕМЫ ЭКСПРЕССИИ КЛОНИРОВАННОГО
ГЕНА, СПОСОБНОЙ ОБЕСПЕЧИТЬ МАКСИМАЛЬНЫЙ ВЫХОД И
ФУНКЦИОНАЛЬНУЮ ПОЛНОЦЕННОСТЬ ПРОДУКТА
Полученные рекомбинантные молекулы переносятся в определенные группы клеток, которые должны обеспечить экспрессию этих генов, то есть синтез соответствующих белков в количествах, экономически рентабельных по сравнению с обычной технологией их производства.
Обычно для данной цели используют бактериальные или дрожжевые культуры клеток, а также системы экспрессии на основе эукариотических клеток.
Из бактериальных клеток наиболее изученной в молекулярно-генетическом отношении является грамотрицательная бактерия Escherichia coli, поэтому для нее можно с наибольшей определенностью планировать генноинженерные конструкции. Однако E. coli слабо освоена промышленностью. Кроме того, она относится к условно-патогенным для человека микроорганизмам, что может создать трудности при получении на ее основе фармацевтических препаратов.
Отмеченные недостатки E. coli легко преодолеваются при конструировании методами генной инженерии штаммов-продуцентов на основе клеток Bacillus subtilis. Данная почвенная бактерия безопасна для человека и животных и прекрасно освоена микробиологической промышленностью. Бактерия B. subtilis по степени изученности следует за E. coli. Важное отличие ее от E. coli - способность эффективно секретировать во внешнюю среду целый ряд белков, поэтому особенно интересны работы по созданию штаммов-продуцентов B. subtilis, секретирующих чужеродные белки из клеток. Однако данная бактерия имеет свои недостатки: рекомбинантные плазмиды в B. subtilis характеризуются нестабильностью, выражающейся в перестройках и делециях ДНК; бациллы секретируют в культуральную среду большое количество протеаз, что существенно усложняет вопрос максимализации генноинженерного получения целевого белка на основе бацилл-продуцентов.
Среди эукариотических микроорганизмов наиболее изученным является низший эукариот Saccharomyces cerevisiae. Одно из преимуществ S. cerevisiae как экспериментальной системы - простота и надежность ее генетического анализа. В клетках дрожжей имеется ферментативная система гликозилирования белков, которая обеспечивает возможность синтеза в них полноценных белков высших эукариот. Аналогичных систем процессинга белков в бактериальных клетках нет. Многие штаммы дрожжей освоены микробиологической промышленностью. Доказана их безвредность для человека и животных. Именно на S. cerevisiae создан первый штамм-продуцент поверхностного антигена вируса гепатита Б, позволивший получить и испытать вакцину против данного вирусного заболевания человека.
С появлением генной инженерии внимание многих исследователей привлекла система культивируемых клеток животных. Особый интерес к культурам клеток животных стал проявляться после обнаружения того, что часть эукариотических генов раздроблена и лишь в системе клеток высших эукариот можно достичь правильной экспрессии таких генов. Кроме того, многие белки животных и их вирусов синтезируются первоначально в виде более высокомолекулярных предшественников, которые в результате специфического протеолитического процессинга переходят в так называемую зрелую форму. Такой процессинг данных белков, по-видимому, можно ожидать лишь в системе клеток животных. Все это убедительно доказывает важность разработки экспрессирующей системы на основе клеток животных.
Реферат опубликован: 15/06/2005 (11529 прочтено)