Страница: 2/2
У миноги эффекты Бора и Хэлдена ориентированы на транспорт CO2. Эффект Хэлдена в интактных эритроцитах миноги настолько велик, что внутриэритроцитарный pH венозной крови выше, чем артериальной. Роль эффекта Хэлдена в транспорте CO2 кровью миноги становится ясной, если рассмотреть механизм этого транспорта. Вследствие относительной непроницаемости мембраны эритроцитов миноги для бикарбоната и других кислотно-основных эквивалентов, карбоангидраза, содержащаяся в эритроците недоступна для бикарбоната плазмы. Вследствие этого только небольшой процент бикарбоната плазмы может быть превращен в CO2 в некатализируемой реакции дегидратации, когда кровь проходит через жабры, потому, что полупериод некатализируемой реакции равен 1 минуте при 15 °С, а время пребывания крови в жабрах всего несколько секунд. Таким образом, большинство удаляемого из крови CO2 в жабрах приходит из эритроцитарных бикарбонатных запасов и эффективность транспорта CO2 зависит от того, какое его количество эритроциты смогут загрузить в тканях и выгрузить в жабрах. Главные факторы, влияющие на это: изменение давления CO2 в кровообращении и кривые диссоциации CO2. На основании кривых диссоциации CO2 для деоксигенированных и оксигенированных эритроцитов Lampetra fluviatilis понятно, что более 80 % уменьшения содержания CO2 в эритроцитах может быть приписано к зависящему от оксигенации уменьшению содержания CO2 и менее чем 20 %-к уменьшению давления CO2. Оксигенация не влияет на содержание CO2 в плазме. Зависящее от оксигенации уменьшение внутриэритроцитарного содержания CO2 полностью объясняется зависящим от оксигенации уменьшением внуриэритроцитарного pH, вызванным эффектом Хэлдана. Таким образом, основываясь на данных in vitro можно сказать, что эффекты Бора и Хэлдана определяюще влияют на транспорт CO2 кровью миноги. Хотя данные in vivo немного отличны, они так же показывают ведущую роль эритроцитов миноги в экскреции CO2 в сравнении: только 15 % выводимого CO2 у радужной форели происходит из эритроцитов.
Из-за высокого венозного внутриэритроцитарного pH (на 0.15-0.2 ед. выше, чем в артериальной) у Petromuzon marinus CO2 эффективно загружается в эритроцитарные запасы, в основном в виде бикарбоната. Вследствие уменьшения давления CO2 крови в жабрах, а так же, вследствие уменьшения внутриэритроцитарного pH из-за высвобождения протонов благодаря оксигенации гемоглобина, CO2 интенсивно выгружается в жабрах. Таким образом, уменьшение общего содержания CO2 при прохождении крови через жабры у миноги подобно таковому у активных костных рыб, таких как радужная форель, за исключением того факта, что бикарбонат плазмы у миноги играет очень ограниченную роль в выведении CO2 из-за медленного проникновения бикарбоната через мембрану.
Влияние концентрации гемоглобина на его функционирование
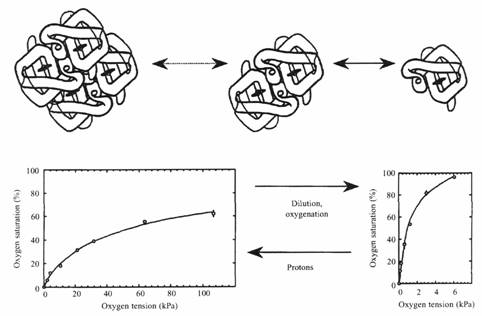
В разбавленных растворах сродство к кислороду гемоглобина миноги заметно зависит от его концентрации. Увеличение концентрации уменьшает сродство благодаря возрастанию процента низкосродственных олигомеров. Так как гемоглобин в интактных эритроцитах показывает кооперативность и эффект Бора, вероятно, реакции ассоциации\диссоциации гемоглобина протекают также и в эритроците. Если это имеет место, то сродство гемоглобина к кислороду должно зависеть от изменения клеточного объема (которое влияет на внутриклеточную концентрацию гемоглобина). Эта зависимость была проверена экспериментально путем осмотического сжатия клеток и последующего измерения P50. Результаты показали уменьшение сродства с уменьшением объема клетки. Кроме этого уменьшения сжатые эритроциты обычно имеют более высокий pH, чем эритроциты с нормальным объемом. Таким образом, концентрационные изменения оказываются способными вызывать изменения в агрегации молекул гемоглобина и, следовательно, сродства гемоглобина к кислороду. Пока не ясно, являются ли подобные изменения объема частью нормальной физиологической регуляцией функции гемоглобина, однако данные об адаптировавшихся к гипоксии Lampetra fluviatilis показывают, что средняя концентрация гемоглобина в эритроците становится ниже, после недели адаптации к гипоксии, чем при нормоксии.
Ограничение буферных свойств гемоглобина вследствие низкой проницаемости мембраны эритроцитов миноги к кислотным эквивалентам.
Заключение
Функционирование гемоглобина в эритроцитах миноги предлагает уникальное среди позвоночных решение газового транспорта. Кооперативность связывания кислорода и влияние протонов на сродство гемоглобина к кислороду проявляются у миноги из-за реакции диссоциации\асоциации молекул гемоглобина. Вследствие низкой проницаемости мембраны эритроцитов для бикарбоната только небольшая часть бикарбоната плазмы может быть дегидрирована до СO2 в течение пребывания крови в жабрах. Однако это ограничение преодолевается высоким внутриклеточным pH и заметными изменениями pH, связанными с оксигенацией. Таким образом, эффективность экскреции CO2 увеличивается.
Хотя транспорт О2 и CO2 кровью миноги так же эффективен, как у активных костных рыб, забуферивание внеклеточных кислотных зарядов неэффективно (из-за непроницаемости мембраны эритроцитов для кислотных эквивалентов). Как следствие: внеклеточные кислотные заряды вызывают заметные колебания pH плазмы у миног.
Таким образом, главное преимущество, полученное, вследствие эволюции анионного обмена, связано не с газовым транспортом, а с буферной системой крови.
Список используемой литературы.
1. Марри, Р., Греннер, Д., Мейес, П., Родуэлл, В., (1993) Биохимия человека ., Москва. изд. «Мир»
2. Шмидт, Р., Тевс, Г., (1996) Физиология человека., Москва. изд. «Мир»
3. Коржуев П. А. (1964) Гемоглобин: сравнительная физиология и биохимия., Москва. изд. «Наука»
4. Nikinmaa, M., Airaksinen, S., Virkki, L. V., (1995) Haemoglobin function in intact lamprey erythrocytes: interactions with membrane function in the regulation of gas transport and acid-base balance. J. Exp. Biol.
5. Ferguson, R. A. Sehdev, N. Bagatto B. Tufts B. L. (1992) In vitro interactions betveen oxygen and carbon dioxide transport in the blood of the sea lampry (Petromyzon Marinus)., J. Exp. Biol.
Реферат опубликован: 16/06/2005 (5287 прочтено)